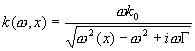|
| |||||||||
|
Renata
Sisto1, Arturo Moleti2 1
Dipartimento Igiene del Lavoro, ISPESL 2
Dipartimento di Fisica, Università di Roma Tor Vergata Sommario
La relazione
fra latenza e frequenza delle emissioni otoacustiche è strettamente legata non
solo alla posizione cocleare dei siti tonotopici, ma anche alla curva di tuning
che esprime l’andamento medio del fattore di merito delle risonanze cocleari in
funzione della frequenza. In questo lavoro presentiamo un modello della
trasmissione cocleare che permette di calcolare la latenza delle emissioni
otoacustiche in funzione della curva di tuning e lo utilizziamo per stimare,
dal confronto con dati sperimentali di latenza, la curva di tuning in adulti e
neonati. Abbiamo inoltre utilizzato i modelli di generazione delle emissioni
spontanee (SOAE) di Zweig e Shera, 1995 e Talmadge et al., 1998, per ricavare
una relazione indipendente fra la latenza e la spaziatura minima delle SOAE, e
abbiamo verificato sui dati sperimentali la correttezza di tali relazioni.
1. Introduzione
Le emissioni otoacustiche (OAEs)
sono segnali acustici misurabili nel canale uditivo esterno sia in risposta a
uno stimolo esterno (emissioni evocate, EOAEs), sia in assenza di stimoli
(emissioni spontanee, SOAEs). Prodotte dai meccanismi di amplificazione attiva
risonante distribuiti lungo la membrana cocleare, costituiscono un importante
mezzo di indagine della funzionalità dell’orecchio interno, ampiamente utilizzato
sia in ambito clinico che nella ricerca fisiologica di base. In ambito clinico,
le emissioni evocate da stimolo transiente (TEOAEs) forniscono semplici criteri
“pass-fail” per lo screening neonatale, mentre per quello che riguarda la
ricerca fisiologica di base, tutte le tecniche sperimentali disponibili (per
una review vedi Probst et al., 1991) sono utilizzate coerentemente insieme ai
modelli teorici per studiare in dettaglio la risposta dell’orecchio. La relazione fra la latenza
delle emissioni otoacustiche e la loro frequenza è stata studiata
sperimentalmente (Tognola et al., 1997, Sisto e Moleti, 2002), e mostra
chiaramente che la latenza sperimentale è ben rappresentata da una semplice
legge di potenza della frequenza. D'altro canto è ben noto che la relazione fra
la frequenza e la posizione del corrispondente sito tonotopico lungo la
membrana cocleare è logaritmica, ed è espressa dalla mappa di Greenwood (1990).
Il confronto fra queste due relazioni empiriche permette di mettere alla prova
modelli teorici di trasmissione del suono lungo la membrana cocleare (Furst et
al., 1988, Talmadge et al., 1998) basati su un formalismo matematico in cui la
membrana cocleare è rappresentata come una linea di trasmissione localmente
risonante a frequenze decrescenti in funzione della distanza secondo la mappa
di Greenwood. In tali modelli la velocità di trasmissione lungo la membrana
cocleare del segnale acustico a una data frequenza è la velocità di gruppo,
determinata dalla relazione che lega localmente il vettore d'onda alla
frequenza. Differenti ipotesi possono essere fatte sulla forma funzionale di
tale relazione. In particolare, una classe importante di modelli teorici e'
basata sull’ipotesi di invarianza di scala, ed e' facile dimostrare che sotto
quest’ipotesi la relazione attesa fra frequenza e latenza cocleare delle
emissioni otoacustiche e' una legge di potenza con esponente –1 (Talmadge et
al., 1998). L'importanza di quest’ipotesi è data dal fatto che, negli stessi
modelli teorici, essa è legata ad un'interpretazione della generazione delle
emissioni otoacustiche che spiegherebbe un aspetto importante della
fenomenologia: la quasiperiodicità delle emissioni spontanee. Sperimentalmente
si osserva invece una relazione tra latenza e frequenza che è in buon accordo
con una legge di potenza, ma con esponente tra –0.4 e –0.8 a seconda degli
autori e delle tecniche utilizzate. D'altra parte, una rottura dell'invarianza
di scala è suggerita anche da evidenze fenomenologiche: 1) a bassa frequenza la
mappa empirica di Greenwood non è esattamente logaritmica e 2) la curva di
tuning in funzione della frequenza ottenuta da misure psicoacustiche (Moore,
1978) suggerisce che il fattore di merito delle risonanze cocleari non sia
costante, ma cresca con la frequenza. Entrambe queste osservazioni implicano
violazioni dell’invarianza di scala. Un'altra possibile spiegazione si ottiene
ammettendo una dipendenza esplicita dalla posizione cocleare dei parametri di
impedenza parallela della lineari trasmissione equivalente. Un’adeguata
parametrizzazione della violazione dell'invarianza di scala associata a queste
cause e la soluzione delle equazioni della linea di trasmissione equivalente
hanno mostrato (Sisto e Moleti, 2002) che è possibile riconciliare i modelli
teorici con le osservazioni sperimentali ipotizzando variazioni del tuning con
la frequenza e/o una dipendenza dell'impedenza parallela dalla posizione
cocleare perfettamente compatibili con le attuali conoscenze della fisiologia
cocleare. In questo lavoro, partendo dai citati risultati, si suggerisce la
possibilità di misurare la curva media di tuning a partire da misure di latenza
delle emissioni otoacustiche, utilizzando eventualmente anche l’informazione
fornita dalla misura della separazione minima fra le SOAE. 2. Materiali e Metodi
In questo lavoro abbiamo analizzato dati di TEOAE e
SSOAE di 94 orecchie di adulti sani e di 22 orecchie di neonati sani, allo
scopo di determinare la relazione tra latenza e frequenza e quella fra
spaziatura minima delle SOAE e frequenza. Due diverse tecniche, descritte
dettagliatamente in Sisto e Moleti (2002), sono state utilizzate per la stima
della latenza, una basata su un’analisi wavelet tempo-frequenza realizzata su
bande di un’ottava, e una basata sull’individuazione, nella forma d’onda della
componente wavelet relativa a una determinata ottava, del picco di risposta
associato ad una particolare riga di frequenza nota, perché osservata anche
come emissione spontanea nello spettro delle SSOAE dello stesso orecchio.
Questa seconda tecnica è ovviamente applicabile solo a un sottoinsieme di casi
in cui la struttura dello spettro delle SOAE è tale da permettere una
individuazione non ambigua delle righe spettrali. La prima tecnica si presta
all’analisi automatica di grandi quantità di dati, ed è stata utilmente
impiegata (Sisto e Moleti, 2002) per valutare differenze statisticamente
significative fra le latenze di soggetti sani e ipoacusici, o anche fra
soggetti sani esposti e non esposti a rumore (Lucertini et al., 2002). Si
tratta però di una tecnica affetta da errori sistematici, che, anche se non
precludono la possibilità di utilizzare le latenze misurate come indicatori da
confrontare fra popolazioni per scopi clinici, possono alterare la dipendenza
della latenza dalla frequenza in modo significativo. Poiché molte delle
proprietà dei modelli cocleari che stiamo cercando di verificare dipendono dai
dettagli di relazioni funzionali come quella che lega la latenza alla
frequenza, utilizzeremo nel seguito il sottoinsieme di dati ricavati dalla seconda
tecnica. 3. Modelli teorici
La latenza delle emissioni
otoacustiche è essenzialmente dovuta al tempo di percorrenza del segnale
acustico lungo la membrana cocleare fino al sito tonotopico corrispondente alla
frequenza in esame e indietro fino alla finestra ovale. E’ possibile tener
conto anche del piccolo (ma non trascurabile) contributo dovuto alla
propagazione del suono nell’orecchio esterno e medio. Poiché l’elemento
dispersivo del sistema uditivo è localizzato nella coclea, è qui che avviene una
dispersione temporale delle diverse componenti in frequenza, che si ripercuote
alla fine in una relazione sperimentalmente misurabile fra latenza e frequenza
delle emissioni otoacustiche. La mappa di Greenwood (1990), che indica la
posizione lungo la membrana cocleare del sito associato a ogni frequenza,
mostra che i segnali di alta frequenza risuonano in siti vicini all’inizio
della membrana (base), mentre quelli di frequenza più bassa risuonano in fondo
alla membrana (apice). Qualitativamente è quindi intuibile che la latenza sia
una funzione decrescente della frequenza, semplicemente perché le componenti a
più bassa frequenza devono percorrere una lunghezza maggiore della membrana
cocleare prima di essere riflesse indietro e venire alla fine rivelate come
emissioni otoacustiche nell’orecchio esterno. Per ottenere una previsione
quantitativa è necessario evidentemente un modello della trasmissione del suono
lungo la membrana cocleare. Un modello utilizzato da molti autori (Furst et
al., 1988, Talmadge et al., 1998, Sisto e Moleti, 2002) con importanti
varianti, che non andiamo qui a discutere, utilizza il formalismo della linea
di trasmissione elettrica equivalente, in cui il valore dei parametri di
impedenza della linea sono associati ad analoghe proprietà “meccaniche” del
sistema reale, che in realtà sono solo in parte puramente meccaniche, essendo
in parte legate al sistema di feedback elettro-meccanico operante nella coclea.
Utilizziamo qui un semplice modello a linea di
trasmissione utilizzato in Sisto e Moleti (2002). La linea è localmente
risonante alla frequenza w(x) nel
punto x della coclea. Risolvendo
l’equazione nel limite WKB si ottiene una relazione fra il vettore d’onda e la
frequenza, che è ovviamente funzione anche di x: E’ possibile quindi calcolare, in funzione di x, la velocità di trasmissione lungo la
membrana cocleare per un pacchetto d’onda di frequenza w, come velocità di gruppo associata alla (1): Una stima della latenza cocleare è quindi data
dall’integrale: In prossimità della risonanza la velocità di gruppo
diminuisce rapidamente, in funzione del Q
della risonanza, su una estensione spaziale che è invece inversamente
proporzionale al Q. E’ quindi
possibile calcolare separatamente il contributo alla latenza dovuto al percorso
vicino alla risonanza e quello del percorso lontano dalla risonanza. In
particolare abbiamo mostrato (Sisto e Moleti, 2002) che il contributo della
risonanza (entro Dx = 2G/w da x(w)) alla latenza è indipendente dal Q e mostra un tipico andamento come w-1, mentre il contributo
lontano dalla risonanza (fino a x(w)-2G/w),
grazie all’approssimata validità dell’invarianza di scala è facilmente
calcolabile in modo analitico: dove: mentre b e’ una costante, dell’ordine di 0.3 ms, che
puo’ essere trascurata in prima approssimazione. Questa relazione permette di ricavare informazioni sulla
curva di tuning a partire da misure sperimentali dalla latenza delle emissioni
otoacustiche texp(w). Si
ricava infatti: D’altra parte, i modelli cocleari delle SOAE basati sulla
riflessione coerente alla Bragg da disomogeneità cocleari in prossimità della
risonanza (Zweig e Shera, 1995; Talmadge et al, 1998) prevedono che la
separazione spaziale minima tra i siti cocleari di origine delle SOAE sia data
dalla condizione: dove dove g e’ quasi indipendente da Q, e vale circa 0.8. La mappa di
Greenwood permette di tradurre l’Eq.(8) in una relazione per la separazione in
frequenza delle SOAE: Abbiamo
quindi una seconda relazione, indipendente dall’Eq.(7), che ci permette di
valutare la curva di tuning partendo in questo caso da misure sperimentali
della separazione minima fra le SOAE:
Le
Eqq.(7) e (11) implicano inoltre una relazione fra latenza e separazione minima
delle SOAE, che deve essere verificata se il modello è coerente: 4. Risultati e discussione
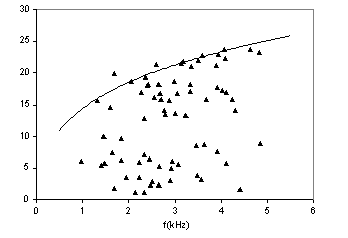
In Fig.1 sono mostrati i valori sperimentali della
latenza in funzione della frequenza, per adulti e neonati e i relativi fit a
una semplice legge di potenza. In Fig.2 sono mostrate le corrispondenti curve
di tuning Q(f), ricavate dall’Eq.(7).
In Fig.3 mostriamo invece l’andamento della separazione fra SOAE in funzione
della frequenza. La linea continua non è un fit ai dati, ma la predizione sulla
separazione minima fornita dall’Eq.(12) a partire dalla misura sperimentale
della latenza. Il buon accordo testimonia della coerenza interna del modello. I
dati sulla latenza di adulti e neonati mostrano una latenza significativamente
maggiore in questi ultimi. Come mostrato in Fig.2, questo corrisponde ad una
ancora più significativa differenza fra le curve di tuning dedotte in base al
nostro modello per adulti e neonati. Il tuning dei neonati risulta
significativamente maggiore di quello degli adulti, con interessanti
implicazioni sulla sensibilità dei neonati al rumore. La solidità di questo
risultato è confermata dalla pressoché perfetta corrispondenza, mostrata in
Fig.3, tra i valori sperimentali della separazione minima fra SOAE dei neonati
e la curva calcolata in base all’Eq.(12) a partire dalla misura della latenza.
Questo significa che avremmo ottenuto lo stessa stima del Q per i neonati a partire da una misura indipendente da quella
della latenza, il che è inoltre una prova della coerenza interna della teoria
utilizzata.
Figura 2
Figura 3 5. Conclusioni Misure di latenza delle OAE e di separazione minima fra
SOAE sono state utilizzate in adulti e neonati per valutarne le curve medie di
tuning, in base a un modello teorico della trasmissione acustica lungo la
coclea. Il Q dei neonati è risultato
significativamente maggiore di quello degli adulti, e il modello ha mostrato un
elevato grado di coerenza interna. Bibliografia
1. Furst, M., and Lapid, M. (1988), J. Acoust. Soc. Am., 84,
222-229.
2. Greenwood, D. D. (1990),
J. Acoust. Soc. Am. 87, 2592-2605. 3. Lucertini M., Moleti A., Sisto R. (2002), J. Acoust. Soc. Am. 111, 972-978. 4. Moore B.C., (1978),
J. Acoust. Soc. Am. 63, 524-532. 5. Probst, R., Lonsbury-Martin, B.L., and
Martin, G.K. (1991), J. Acoust. Soc. Am. 89,
2027-2067 6. Sisto R., Moleti A. (2002), J. Acoust. Soc. Am. 111,
297-308. 7. Talmadge, C.L., Tubis, A., Long, G.R., Piskorski, P. (1998), J. Acoust. Soc. Am. 104,
1517-1543. 8. Tognola, G., Grandori, F., Ravazzani, P. (1997), Hear. Res. 106, 112-122. 9. Zweig G., Shera C.A. (1995), J. Acoust. Soc. Am.
98, 2018-2047. |