 |
| |||
| Physiopathological significance of distortion-product otoacoustic emissions at 2f1-f2 produced by paper published in J.Acoust.Soc.Am. 2003, 113, 430-441
1. OVERVIEW Akin to the other classes of otoacoustic emissions (OAE), distortion product OAEs or DPOAEs at 2f1-f2 in response to pure-tone stimuli at f1 and f2 are viewed as a reliable tool (and moreover, a frequency-specific one) for tracking outer hair cell (OHC) dysfunctions. However, when DPOAEs are used for screening, that is mainly in a pass / fail paradigm, it is widely acknowledged that their "audiometric" reliability is no longer granted for stimulus levels exceeding 60-70 dB SPL. This is because residual DPOAEs may well exist in spite of patent cochlear dysfunction, as observed in numerous examples involving short-term cochlear ischemia or furosemide injection (Mills, 1997, Mom et al., 1997). Thus it is not advisable to screen for OHC-related hearing deficit with DPOAEs elicited by stimuli > 60 dB SPL (from now on we'll call them high-level DPOAEs, as opposed to low-level DPOAEs elicited by stimuli below 60 dB SPL). Such an empirically-based caveat raises concerns for three reasons, a fundamental one and two practical ones. The 1st practical reason is that DPOAEs in humans are not very high-level signals and that their signal-to-noise ratio is more comfortable when stimuli > 60 dB SPL are used. The 2nd reason is that many default setups of commercially available equipments start with 60, and even 70 dB SPL primary levels (likely on account of the 1st reason: it is not a good advertisement for a product if it turns out to be unable to detect DPOAEs in an otherwise fully normal ear). The 3rd, fundamental reason is that the fact of obtaining a surprising result, such as the presence of residual high-level DPOAEs from a "dead" cochlea, does not constitute a sufficient reason to discard it without further afterthought. Although their being unsuitable for screening or audiometric purposes is clearly established, high-level DPOAEs might have some physiological significance that could well lead to suggest another interesting use for them. This is not purely wishful thinking: the very origin of OAEs and of distortion has always seemed tightly correlated with the intimate, functional or structural properties of sensory cells in the organ of Corti. If, instead of reflecting "active" cochlear motion, high-level DPOAEs represented another, "passive" modality, as suggested for example by Mills (1997), it would be a good reason for eventually dismissing them. Mills' model is an elegant attempt at reconciling the high vulnerability of low-level DPOAEs to cochlear insults with the robustness of high-level DPOAEs to some types of cochlear damage. It posits that DPOAEs are the vector sum of two components, an "active", slow-growing and vulnerable one, and a "passive" one, virtually absent at low levels, but which would grow so fast as to eventually dominate the whole DPOAE signal at high levels. As a result, when pathology kills the low-level "active" component, the overall DPOAE signal would vanish at low levels but conversely would vary little at high levels. In the intermediate range, in normal cochleas, the "active" and "passive" components might happen to be equal in size and just out of phase, thereby canceling each other, hence the notch often observed in DPOAE growth functions around 60 dB SPL. It is noteworthy that recent publications by Lukashkin and Russell (1999) have taken exception to this interpretation and explained the notch by a simpler, one-source model. David Mills' model still assumes that even "passive" DPOAEs can be produced by OHCs. Only, these OHCs would extend far basalward to the place tuned to f1and f2 and their motion would only reflect the passive cochlear macromechanics, known to be unaffected by cochlear pathology. Although "passive" DPOAEs would still have to do with OHCs, their functional interest would be restricted. More pessimistic interpretations than Mills' even exist wherein "passive" DPOAE sources would be artifacts from the basilar membrane, inner hair or support cells, thus losing any interest. In the present paper, the authors have adopted an alternative standpoint wherein regardless of stimulus level, cubic DPOAEs are produced by N (probably OHC-borne) nonlinear elements forming a nonlinear "black box". Instead of calling them "active" or "passive" depending on which modality the cochlea was assumed to operate on, they considered that the output of any nonlinear system must be easily predictable from the knowledge of its input I, of the number of its elements N, and that a coefficient accounting for the exact shape of the nonlinearity had to be included, for example by introducing a function F3 (3 because the DPOAE of interest is a cubic one) of the operating point o of the nonlinearity. Thus the authors set out to test the following relationship: (again, I to the power 3 because the DPOAE of interest at 2f1-f2 is a cubic one). 2. MAIN RESULTS 2.1. CD1 mice, genetic cochlear impairment
The interest of this strain (Shone et al., 1991; Le Calvez et al., 1998a), is that the hearing losses and OHC coytocochleograms often show a very sharp boundary somewhere between an intact apex and a damaged base, the position of which depends on age. In the present work, the DPgram patterns (i.e., the plots of DPOAE levels against frequency f2) paralleled those of ABR thresholds against frequency at all primary levels. In ears with steep ski-sloped audiograms, similarly steep DPgrams were found regardless of stimulus level even when > 70 dB SPL. Likewise, highly significant correlations between DPOAE levels and ABR thresholds at f2 were found even at the highest tested primary levels, that is, 75 or 80 dB SPL. Overall, DPOAE levels decreased with ABR thresholds, and instrumental distortion was reached when ABR threshold elevation exceeded about 70 dB nHL.
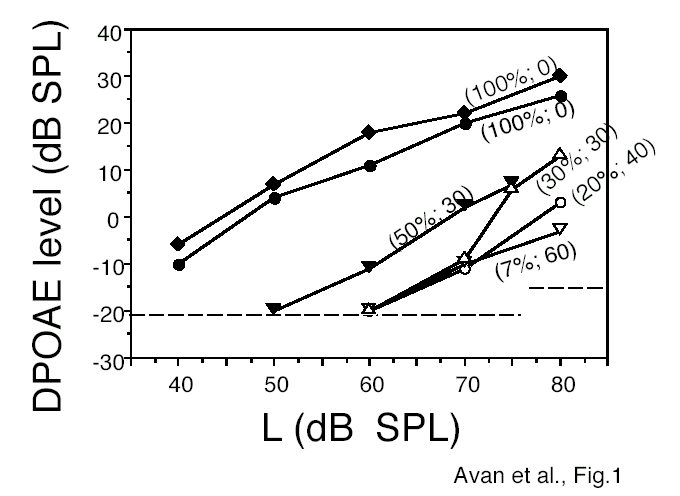
Input-output DPOAE curves are plotted in Fig.1. Percentages of remaining OHCs, all rows included, and ABR thresholds in dB nHL are printed between brackets along with every curve. While the two top curves correspond to intact OHCs in two cochleae with normal ABR thresholds at 15 kHz, and DPOAEs still present at 40 dB SPL, conversely, the two bottom curves exhibit DPOAEs above noise floor only above 70 dB SPL. The corresponding percentages of extant OHCs are the lowest, namely 7 and 20%. ABR thresholds are the most elevated ones, 40 and 60 dB nHL. The remaining plots lay between these two extremes. In summary, when the stimulus level exceeded 70 dB SPL, only the cochlear regions with OHCs produced high-level DPOAEs, in proportion to the residual number of OHCs, as predicted by model (1). No "passive" high-level DPOAE existed when no OHC was present in the area of interest (i.e., that tuned to f2). Using high-level stimuli did not prevent the boundary between healthy and damaged regions to be visible and correctly situated.
Fig.1: DPOAE levels vs. level L of equilevel stimuli in six CD1 ears tested at f2 = 15 kHz. In parentheses: 1st figure = percentage of residual OHCs at the place tuned to f2; 2nd figure = ABR threshold in dB nHL in response to tone-pips of center frequency f = 15 kHz. Dashed horizontal lines: limits of background noise (below 75 dB SPL) or instrumental distortion (at 80 dB SPL). The three data points near the background noise limit may be overestimations of true DPOAE levels.
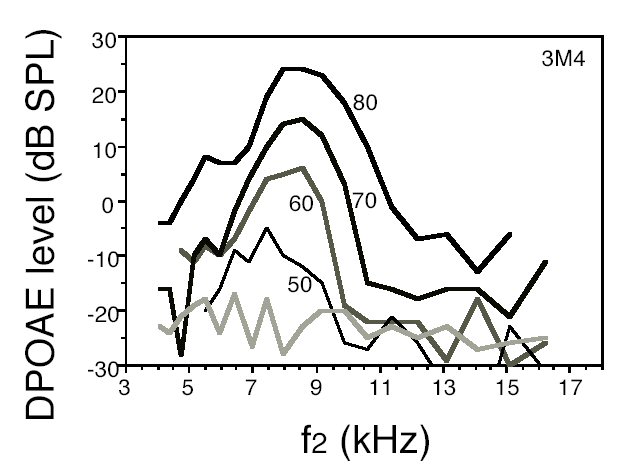
Fig.2: Example of a series of DPgrams collected with equilevel stimuli (80 to 50 dB SPL as indicated by labels on each DPgram) from the right ear of mouse 3M4 (7 weeks of age). Hatched line: average background noise.
2.2. Gerbils and selective cochlear ischemia
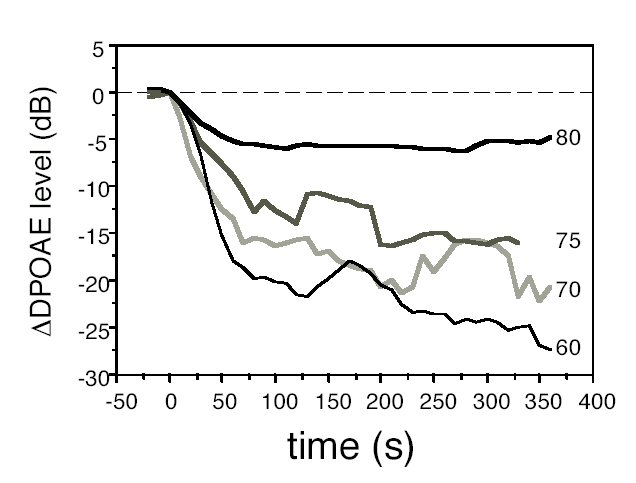
As already described in the literature (Mom et al., 1997), DPOAE level changes with time were monotonic over the first minute of ischemia then a plateau was reached either directly or, in more than half individual ears and below 80 dB SPL, after a level notch followed by a rebound. The plateau lasted until at least 6 min after ischemia onset, after which the pressure by the pipette by which ischemia was effected in the porus acusticus was released and the monitoring of cochlear blood flow and DPOAEs interrupted. The plateau level drops relative to pre-ischemic levels gradually decreased in size with increasing level: on average, from -25 dB at 60 dB SPL to -5 dB at 80 dB SPL. Ischemic growth functions became steeper and more monotonic than before ischemia: the ischemic slope was about 1.8 dB / dB between 60 and 80 dB SPL and approached 3 dB / dB between 75 and 80 dB SPL. Low-level DPOAEs were more "sensitive" to ischemia in that their levels decreased more than high-level DPOAEs, as predicted by the "active / passive" model but also by model (1). On the other hand, the results of monitoring at 80 dB SPL actually showed that high-level DPOAEs reacted slightly faster to the cochlear impairment. This is not in line with what would be expected if such DPOAEs were passive ones. It suggests that they are vulnerable, only, according to model (1), high primary levels tend to make up for the lack of cochlear gain (I is unchanged around 80 dB SPL). Besides, short-duration ischemia in no way affects N: the cochlear function recovered if the cochlear blood flow was let back to normal after the 6 min this experiment lasted. We thus propose that the ischemic DPOAEs come from the same OHC-related structures that produced them normally, because ischemia does not affect their structure and the high-level stimuli drive them into the nonlinear part of their characteristics just the same as in a healthy cochlea. Monitoring with high level stimuli would tell us nothing relative to the audiogram, however, it definitely tells us that OHCs are still there, able to perform at least part of their function (the one that produces DPOAEs) when their input is large enough. 2.3 Gerbils, furosemide vs. cochlear ischemia In this part of the experiment, round-window cochlear microphonics (CM) and DPOAEs were simultaneously recorded in gerbils, either after ischemia followed by reperfusion, or after IV injection of the loop diuretics furosemide. Both impairments were fully reversible after a few minutes, so that the time courses of CM and DPOAE changes could be followed up and compared along the course of growing impairment, then recovery. CM changes were rather similar in both experiments, and in order to offset as much as possible the slight differences observed, the ratio of DPOAE / CM changes was computed and analyzed. It turned out that whereas in furosemide experiments, the ratio DP / CM varied little and remained close to 1 throughout an experiment, in ischemia / reperfusion, it showed wild variations. As N and I were assumed to be very similar in the two experiments, one has to assume that another factor in (1) had to be different. Although it is only a conjecture, F3(o), relating to the exact shape of the nonlinear function at the origin of DPOAEs, could play the appropriate role. SUMMARY In summary, the present results suggest that, provided of course the recording equipment is reliably distortion-free (it is easier to fulfill this condition in small rodents because the volume of their outer ear canal is very small), the persistence of high-level DPOAEs reveals the persistence of functional nonlinear elements in the cochlea. These nonlinear elements could well be hosted by OHC stereocilia bundles (Strelioff and Flock, 1984), for example, they could be the mechanoelectrical transduction channels as their nonlinear characteristics (relating K+ current to deflection) are well acknowledged. The proposed model (1) fits the present experimental results better than the "active / passive" one, and is more parsimonious, as already suggested by the theoretical analysis of Lukashkin and Russell (1999). The results lead to propose that distortion is mainly a mechanical (thus rather "passive") phenomenon, that may persist even though the cochlear gain is turned off, provided the nonlinear elements survive and receive enough mechanical input. Although according to (1), high-level DPOAEs are expected to be poor audiometric indicators (as they have indeed been shown to be), they seem well adapted to assessing the functional integrity of nonlinear elements in OHCs, i.e., presumably their mechanoelectrical transduction channels.
Sample of useful REFERENCES
Bian, L. and Chertoff, M.E. (2001), "Distinguishing cochlear pathophysiology in 4-aminopyridine and furosemide treated ears using a nonlinear systems identification technique," J.Acoust.Soc.Am. 109, 671-685. Flock, A. and Strelioff D. (1984), "Graded and nonlinear mechanical properties of sensory hairs in the mammalian hearing organ," Nature 310, 597-599. Frank, G. and Koessl, M. (1996), "The acoustic two-tone distortions 2f1-f2 and f2-f1 and their possible relations to changes in the operating point of the cochlear amplifier," Hear.Res. 98, 104-115. Le Calvez, S., Avan, P., Gilain, L. and Romand, R. (1998), "CD1 hearing-impaired mice. I: Distortion product otoacoustic emission levels, cochlear function and morphology," Hear.Res. 120, 37-50. Lukashkin, A.N. and Russell, I.J. (1999), "Analysis of the f2 - f1 and 2f1 - f2 distortion components generated by the hair cell mechanoelectrical transducer: Dependence on the amplitudes of the primaries and feedback gain," J.Acoust.Soc.Am. 106, 2661-2668. Mills, D.M. (1997), "Interpretation of distortion product otoacoustic emissions measurements: I. two stimulus tones," J.Acoust.Soc.Am. 102, 413-429. Mom, T., Avan, P., Romand, R. and Gilain, L. (1997), "Monitoring of functional changes after transient ischemia in gerbil cochlea," Brain Res. 751, 20-30. Ruggero, M.A. and Rich, N.C. (1991), "Furosemide alters organ of Corti mechanics: evidence for feedback of outer hair cells upon the basilar membrane," J.Neurosci. 11, 1057-1067. Whitehead, M.L., Lonsbury-Martin, B.L. and Martin, G.K. (1992), "Evidence for two discrete sources of 2f1-f2 distortion-product otoacoustic emission in rabbit: II. Differential physiological vulnerability," J.Acoust.Soc.Am. 92, 2662-2682.
| |||
 |